DETECCI”N DE QTLs PARA DEPOSICI”N DE GRASA Y CRECIMIENTO EN EL CROMOSOMA 4 EN UN CRUCE F2 DE CERDO IB…RICO Y LANDRACE
Clop A . (1) , Pťrez Enciso M. (2) , SŠnchez E. (1), ”vilo C. (3), Coll A. (1), Folch J.M. (1), SŠnchez A.. (1), Estany J. (2), Babot D. (2), Noguera J.L. (2)(1) Unitat de GenŤtica i Millora animals, Fac. de Veterinŗria, Universitat AutÚnoma deBarcelona, 08193 Bellaterra. (2)ņrea de Producciů Animal, Centre UdL-IRTA. E-25198 Lleida. (3)Area de Mejora Genťtica animal, CIT-INIA, 28040 Madrid.
INTRODUCCI”N
La detecciůn de QTLs (Quantitative Trait Loci), regiones del genoma que intervienen en el valor cuantitativo de un determinado carŠcter, ha sido objeto de un creciente interťs en los ķltimos aŮos a partir de la existencia de marcadores moleculares (microsatťlites, AFLPs, etc...) en el genoma de las principales especies domťsticas. En el mapa genťtico porcino existen numerosos microsatťlites bien localizados y altamente polimůrficos (Rohrer et al., 1996).
Se realizů un cruce experimental tipo F2 de cerdos de la raza Ibťrica con cerdas Landrace con el objetivo de detectar marcadores moleculares ligados a caracteres de interťs productivo en porcino. Estas dos poblaciones tienen caracterŪsticas bien diferenciadas en cuanto a la calidad de la carne (IBMap Consortium, 1998 y Serra et al., 1998). El cruce entre individuos fenotŪpicamente muy distanciados incrementa la potencia estadŪstica de este tipo de diseŮo.
En este trabajo presentamos los datos relativos a la detecciůn de QTLs en el cromosoma 4 porcino, con efecto sobre la deposiciůn de grasa y el crecimiento, dos caracteres de importancia para la industria porcina. Paralelamente, se ha realizado con el mismo material animal, un estudio sobre QTLs que afectan a la calidad de la carne en el cromosoma 6 porcino (”vilo et al., 1999).
MATERIAL Y M…TODOS
Animales. Tres machos Ibťricos pertenecientes a la lŪnea Guadyerbas fueron cruzados con 31 hembras Landrace homocigotas dominantes en el diagnůstico genťtico del gen Hal (SŠnchez et al., 1993). La F1 estŠ formada por 6 machos y 73 hembras que han generado 577 individuos F2. El manejo de los animales ha sido el habitual en condiciones comerciales.
En este trabajo presentamos los resultados correspondientes al anŠlisis de 247 F2, agrupados en 33 familias de hermanos. El sacrificio se llevů a cabo en 4 lotes a 175.5 Ī 0.3 dŪas de edad y un peso de canal promedio de 74.7 Ī 0.6 Kg. Los caracteres estudiados fueron el peso de la canal corregido por edad al sacrificio (P.C.), espesor de grasa entre la 3™ y 4™ costilla y a unos 6 cm de la lŪnea media (G2), peso de la grasa dorsal en la mitad izquierda de la canal obtenida en un despiece comercial (P.G.D.), y porcentaje de magro (% magro), estimado a partir de G2 y la profundidad del lomo utilizando la ecuaciůn de regresiůn propuesta por Gispert y Diestre (1994). Estos tres ķltimos caracteres se corrigieron por peso de la canal.
ADN. La extracciůn de ADN genůmico se realizů a partir de sangre. Para la poblaciůn parental se utilizů un protocolo de precipitaciůn salina (Miller et al., 1988). Para la F1 y la F2 se utilizů un kit comercial (Boehringer Mannheim) basado, tambiťn, en una precipitaciůn salina.
Genotipados. Las reacciones de amplificaciůn de los microsatťlites se realizaron en un equipo automatizado de PCR ABI PRISM 877 Integrated Thermal Cycler de Perkin Elmer. Los productos de PCR con cebadores marcados fluorescentemente se analizaron en un equipo de electroforesis capilar y detecciůn fluorescente (ABI PRISM 310 Genetic Analyzer). En la elecciůn de los microsatťlites se tuvo en cuenta la posiciůn de ťstos en el genoma y su variabilidad, estimada mediante el Ūndice de informatividad de Ron (Ie) (Ron et al., 1995), que fue estimada a partir de una muestra de la poblaciůn parental (Clop et al., 1998). Para el cromosoma 4, se seleccionaron 7 microsatťlites. La construcciůn posterior del mapa se realizů utilizando el programa Cri-Map (Green et al., 1990) .
AnŠlisis estadŪstico. El modelo para el anŠlisis de QTLs incluye el sexo, lote y genotipo del QTL como efectos fijos, asŪ como peso canal como covariable, excepto en el anŠlisis del peso canal que se corrigiů por la edad al sacrificio. El anŠlisis se realizů mediante mapeo de intervalos con regresiůn (Haley et al., 1994), que permite incluir la informaciůn de todos los marcadores cuando los genotipos no son informativos. Se asume que existe un QTL con alelos alternativos fijados en cada raza con valores a y –a para los homocigotos de cada raza, y d para los heterocigotos. El modelo que incluye el QTL se compara con el modelo sin QTL cada cM mediante un test F clŠsico.
RESULTADOS Y DISCUSI”N
Mapa de ligamiento: El mapa resultante de nuestros datos se asemeja al descrito por otros autores (Rohrer et al., 1996) y cubre 121 cM. La longitud del cromosoma 4 es de 144 cM en las hembras y 127,8 cM en los machos. La informatividad de los microsatťlites, calculada sobre los individuos de la F2 (contenido informativo de la F2) coincide con la estimada mediante el Ūndice de Ron (Ie) a partir de una muestra de animales de la poblaciůn parental (Tabla 1). Por lo tanto puede tomarse como criterio para seleccionar microsatťlites segķn su nivel de informatividad.
Tabla 1
Microsatťlite |
Posiciůn Rorher et al. (1996) |
Posiciůn (cM) presente trabajo |
Ie |
Contenido Informativo F2 presente trabajo |
SW2404 |
0 |
0 |
0.57 |
0.510 |
S0301 |
27 |
36 |
0.84 |
0.805 |
S0001 |
42 |
55 |
0.89 |
0.844 |
SW839 |
63 |
65 |
0.96 |
0.992 |
S0214 |
81 |
83 |
0.96 |
0.998 |
SW445 |
106 |
105 |
1.00 |
0.948 |
S0097 |
120 |
121 |
0.86 |
0.773 |
La informatividad fue muy alta en 6 de los 7 microsatťlites, siendo SW2404 el de menor contenido informativo.
AnŠlisis de QTLs. El valor aditivo a y el dominante d se calcularon sobre la mejor estima de la posiciůn del QTL obtenida mediante el test F clŠsico (Tabla 2). El valor mŪnimo de significaciůn de F en otros trabajos similares a ťste, se sitķa alrededor de 8 (Andersson et al, 1994; Walling et al, 1998).
Tabla 2
CarŠcter |
Posiciůn (cM) |
F mŠx. |
a |
d |
P.C. |
77 |
5.682 |
-2.334Ī0.859 |
1.805Ī0.129 |
G2 |
71 |
11.337 |
2.453Ī0.569 |
-1.073Ī0.862 |
P. G.D. |
70 |
15.138 |
0.24Ī0.046 |
-0.046Ī6.98 |
% magro |
71 |
14.439 |
-2.344Ī0.471 |
0.813Ī0.715 |
El peso canal es el ķnico carŠcter estudiado que no presenta un valor F mŠximo significativo. Los restantes factores superan claramente el mŪnimo significativo.
El alelo ibťrico implica disminuciůn de P.C. y % magro, mientras que G2 y P.G.D. se ven incrementadas cuando la variante ibťrica se encuentra presente. Esto concuerda con que la raza Ibťrica es mucho mŠs grasa que la Landrace. En la Figura 1 se muestran los valores de F para cada cM del cromosoma 4, para los caracteres medidos.
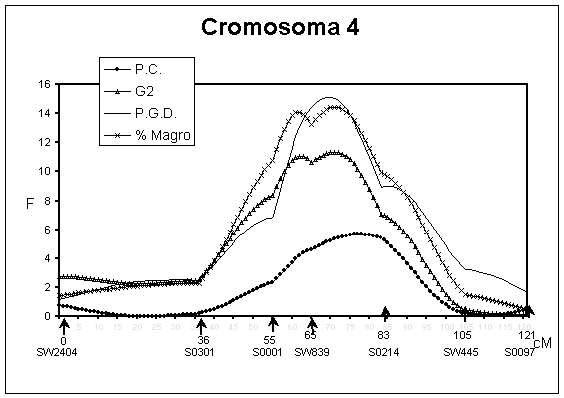
Figura 1
Por la posiciůn cromosůmica en que se ha detectado, resulta probable que se trate de un mismo QTL con efecto sobre P.C., G2, P.G.D., y % magro. Asimismo parece razonable pensar que sea el mismo detectado por Andersson et al. (1994) y a su vez, por Walling et al. (1998) en otras poblaciones. No obstante, el valor que adquiere en nuestro estudio el peso canal no es significativo, quizŠs debido a que los alelos no estťn fijados en cada poblaciůn.
No existen evidencias de un efecto directo sobre alguno de estos caracteres en los genes localizados en este cromosoma. Sin embargo, en la regiůn detectada, hay algunos loci estructurales que podrŪan tener cierta influencia sobre ellos. Entre ťstos se encuentra la subunidad beta 1 de la ATPasa 1 (ATP1B1), los factores de transcripciůn OTF, SSC9C8, la beta glicosidasa (GBA) y la subunidad beta de la hormona estimulante del tiroides (TSHB).
AGRADECIMIENTOS
Al SIA Dehesůn del Encinar que facilitů los parentales Guadyerbas. A Pere Borrŗs, Eva Ramells y en general a todo el personal de Nova GenŤtica y del matadero de Copaga por su inestimable cooperaciůn. A AgustŪ Quintan y Meritxell Arquťs por la ayuda en la toma de muestras. A Max Rothschild y al USDA supported US Pig Genome Coordination Project por el generoso envŪo de cebadores A Chris Haley por los programas de anŠlisis y al resto del equipo IBMAP. Trabajo financiado por la CICYT (proyecto AGF 96-2510). ņlex Clop tiene una predoctoral de formaciůn de investigadores concedida por la Universitat AutÚnoma de Barcelona.BIBLIOGRAFÕA
-Andersson L. et al.: Science, vol. 263 1771-1774, 1994.
-Archibald A. et al.: Mammalian Genome, 6. 157-175, 1995.
-Clop A. et al.: IX Reuniůn Nacional de Mejora Genťtica Animal, 1998. Libro de Resķmenes. ITEA.
-Green P. et al.: Documentation for CRI-MAP, version 2.4, 1990.
-Gispert M., Diestre, A.: Techniporc 17:2 1994.
-Haley C. et al.: Genetics 136, 1195-1207 1994.
-Marklund L. et al.: Animal Genetics, 27. 256-269, 1996.
-Miller S. A. et al.: Nucleic Acid Research, 16, n. 3, 1215.
-”vilo et al.: ITEA, 1999 (enviado).
-Ron M. et al.: Animal Genetics, 26. 439-441, 1995.
-Rorher G. et al.: Genome Research, 6:371-391, 1996.
-SŠnchez et al., ITEA 12: 257-259 1993.
-Serra X. et al., Livestock Production Science 56: 215-232, 1998 .
-The IBMAP Consortium: 6th world Congress on genetics applied to livestock production, 1998.
-Walling G. et al.: Animal Genetics 29. 415-424, 1998.