EFECTOS DE LA SELECCION DIVERGENTE POR CAPACIDAD UTERINA SOBRE LA TASA DE OVULACIÆN Y EL PESO EN CONEJO
Argente, M.J*; Santacreu, M.A+; Climent, A+; Blasco, A.+
*Departamento de TecnologÚa Agroalimentaria. Divisiµn de Producciµn Animal. Campus de Orihuela. Ctra de Beniel, Km 3,2. 03312 Orihuela. Alicante.
+
Departamento de Ciencia Animal. Universidad PolitÕcnica de Valencia.Apartado 22012. Camino de Vera, 14. 46071 Valencia.
INTRODUCCION
Christenson et al. (1987) definieron la capacidad uterina como el mÃximo nºmero de fetos que una hembra es capaz de llevar al tÕrmino de la gestaciµn cuando el nºmero de µvulos no es el factor limitante. En conejo, Blasco et al. (1994) proponen estimar la capacidad uterina como el tamaþo de camada de las hembras ovariectomizadas unilateralmente (ULO).
En ratµn, la mejora en el tamaþo de camada ha ido acompaþado de un incremento en la tasa de ovulaciµn y en el peso (Joakimsen y Baker, 1977). El objetivo de este trabajo es el estudio de la relaciµn existente entre el peso y la tasa de ovulaciµn, y analizar si las diferencias encontradas por Argente et al. (1996) en el experimento de selecciµn divergente por capacidad uterina en conejo estuvieron asociadas a diferencias en el peso de la hembra.
MATERIAL Y METODOS
Animales
Los animales pertenecen a un experimento de diez generaciones de selecciµn divergente por capacidad uterina (Argente et al., 1996). A todas las hembras se les extirpµ su ovario izquierdo (hembras ULO) antes de alcanzar la pubertad. A los 12 dÚas de su segunda gestaciµn, todas las hembras fueron pesadas, y se les prÃctico una laparoscopÚa para contabilizar el nºmero de cuerpos lºteos y el nºmero de embriones implantados. A partir de la generaciµn siete, tambiÕn se pesµ la hembra a la monta. Se dispuso de un grupo de hembras intactas contemporÃneas desde la generaciµn quinta.
Caracteres
Los caracteres analizados fueron: CU:capacidad uterina medida como el tamaþo de camada en las hembras ovariectomizadas unilateralmente (ULO). TO:tasa de ovulaciµn. EI:nºmero de embriones implantados. PM:peso de la hembra a la monta en su segunda gestaciµn. PL:peso de la hembra a los 12 dÚas de la segunda gestaciµn. GP:ganancia de peso desde la monta hasta los 12 dÚas de la segunda gestaciµn.
AnÃlisis EstadÚsticos
Las medias por mÚnimos cuadrados se obtuvieron con un modelo que incluÚa los efectos fijos: tratamiento (hembras ULO o intactas), estaciµn, generaciµn, lÚnea jerarquizada a generaciµn, y estado de lactaciµn (lactantes o no lactantes). Las diferencias entre pesos se analizaron con la covariable edad a la monta. Se aþadiµ la covariable peso y la interacciµn entre Õsta y el tratamiento, como esta interacciµn no fue significativa en ningºn caso, se suprimiµ de los anÃlisis.
RESULTADOS Y DISCUSION
La tabla 1 muestra cµmo la ovariectomÚa unilateral no afectµ ni el peso a la monta (PM) y ni el peso a los 12 dÚas de gestaciµn (PL) en las hembras ULO. La ganancia de peso desde la monta hasta los 12 dÚas de gestaciµn fue mayor en las hembras intactas, probablemente debido a que gestaron dos embriones mÃs que las hembras ULO.
La figura 1 muestra la evoluciµn de la tasa de ovulaciµn (TO) y del peso de las hembras (PL) durante las diez primeras generaciones de selecciµn divergente por capacidad uterina. Ni la TO ni el PL parecen verse afectados por la selecciµn por capacidad uterina. Sin embargo en ratµn, las lÚneas seleccionadas por tamaþo de camada, un Úndice o capacidad uterina incrementaron el peso a la monta respecto a la lÚnea control (Van Engelen et al., 1995).
Las correlaciones residuales estimadas entre la TO y los pesos a la monta y a la los doce dÚas de gestaciµn variaron de +0.10 a +0.23. Estos valores son inferiores al +0.54 encontrado en conejas nulÚparas por Molina et al. (1987).
La tabla 2 muestra los resultados de los anÃlisis de regresiµn lineal. El peso de la hembra està relacionado positivamente con la TO. El nºmero de embriones implantados sµlo parece estar relacionado con el peso a los 12 dÚas de gestaciµn. El cociente R2/R2mÃx (Draper y Smith, 1981) muestra como el peso aumenta conforme la TO y el EI se incrementa.
Respecto al efecto de lactaciµn, se ha observado que las hembras que estaban gestantes y lactantes a la vez tenÚan una mayor tasa de ovulaciµn (0.61 µvulos, P<0.01) y un mayor peso a la monta (185 g, P<0.10), aunque esta ºltima diferencia desapareciµ a la implantaciµn. En la bibliografÚa, los resultados encontrados son contradictorios. Foxcroft y Hasmain (1973) observaron un efecto favorable significativo de la lactaciµn sobre la tasa de ovulaciµn. Sin embargo, GarcÚa y PÕrez (1989) y Fortun et al. (1993) no encontraron ningºn efecto significativo del estado de lactaciµn.
Tabla 1. Nºmero de datos (N), medias por mÚnimos cuadrados (MMC), y error estÃndar (EE) en el grupo de hembras ULO e intactas.
N |
ULO |
INTACTAS |
|||||
MMC |
EE |
N |
MMC |
EE |
Sig |
||
TO |
736 |
13.98 |
0.10 |
194 |
14.76 |
0.20 |
** |
EI |
736 |
10.52 |
0.12 |
194 |
12.67 |
0.24 |
** |
PM (kg) |
245 |
4.04 |
0.03 |
97 |
4.08 |
0.05 |
ns |
PL (kg) |
732 |
4.18 |
0.02 |
190 |
4.18 |
0.03 |
ns |
Pmedad (kg) |
245 |
4.03 |
0.03 |
97 |
4.05 |
0.05 |
ns |
Pledad (kg) |
732 |
4.18 |
0.02 |
190 |
4.17 |
0.03 |
ns |
GP (kg) |
180 |
0.14 |
0.02 |
60 |
0.21 |
0.03 |
* |
TO: tasa de ovulaciµn. EI: nºmero de embriones implantados. PM: peso a la monta. PL: peso a los 12 dÚas de la segunda gestaciµn. PMedad y PLedad: peso a la monta y a los 12 dÚas de la segunda gestaciµn corregido por la edad de la hembra. GP: ganancia de peso entre la monta y los 12 dÚas de gestaciµn. **:P<0.01. *:P<0.05. ns:no difieren significativamente.
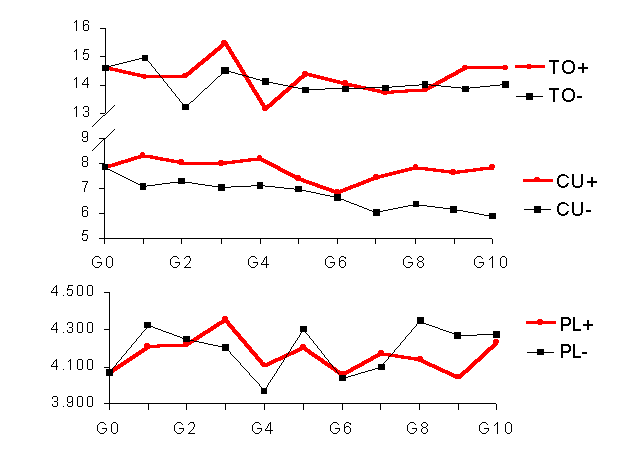
Figura 1. Evoluciµn de las diferencias entre la lÚnea de alta y de baja capacidad uterina a lo largo de las diez primeras generaciones de selecciµn en la capacidad uterina (CU), la tasa de ovulaciµn (TO) y el peso a los 12 dÚas de gestaciµn (PL).
Tabla 2. Coeficiente de regresiµn (b) con su error estÃndar (EE) del peso a la monta (PM) y a los 12 dÚas de gestaciµn (PL) sobre la tasa de ovulaciµn (TO) y nºmero de embriones implantados (EI).
ULO+INTACTAS |
||||||
Y |
X |
N |
b+EE |
R2 |
R2/R2mÃx |
Sig |
PM |
TO |
243 |
0.020+0.011 |
0.10 |
0.60 |
+ |
PL |
TO |
921 |
0.039+0.005 |
0.13 |
0.87 |
** |
PM |
EI |
243 |
0.003+0.009 |
0.09 |
0.61 |
ns |
PL |
EI |
921 |
0.015+0.005 |
0.09 |
0.71 |
** |
Y:variable dependiente. X:covariable. **:P<0.01.+: P<0.10. ns:no difieren significativamente.
REFERENCIAS
Argente, M.J.; Santacreu, M.A.; Climent, A.; Blasco, A.. 1996. 6th World Rabbit
Congress. Toulouse. Vol ll:241-244.
Blasco, A; Argente, M.J; Haley, C.S; Santacreu, M.A. 1994. J. Anim. Sci.
72:3066-3072.
Christenson, R.K; Leymaster, K.A; Young, L.D. 1987. J. Anim. Sci. 67:738-744.
Draper, N.R.; Smith H. 1981. John Wiley&Sons, New York.
Fortun, L.; Prunier, A.; Lebas, F. 1993. J. Anim. Sci. 71:1882-1886.
Foxcroff, G.R.; Hasnain, H. 1973. J. Reprod. Fertil. 33(3):367-377.
Garcia, F.; PÕrez, A. 1989. ITEA, 80:3-10.
Joakimsen, O.; Baker, R.L.1977. Acta Agriculturae Scandinavica, 27:301-318.
Molina, I.; Pla, M.; Garcia, F. 1987. ITEA. 68:61-66.
S.A.S. 1998. SAS User 's Guide: Statistics. SAS Inst. Inc., Cary, NC.
Van Engelen, M.A.J.;Nielsen, M.K.; Ribeiro, E.L.A. 1995. J. Anim. Sci. 73:1948-1953.