Fundadores o ancestros en el análisis genealógico: aplicación a la estirpe Guadyerbas de cerdo ibérico
M. Toro, J. Rodrigáńez, L. Silió y M.C. Rodríguez
Departamento de Mejora Genética y Biotecnología, INIA, Madrid
Introducción
El mantenimiento de la diversidad genética es el objetivo primordial de los programas de conservación. En los últimos ańos ha existido un creciente interés tanto en el desarrollo de técnicas para el análisis genealógico (goteo de genes, por ejemplo) como en proponer parámetros que resuman la evolución de la variabilidad genética. Recientemente Caballero y Toro (1999) han descrito estos parámetros y su interrelación. En este artículo se presenta la evolución de estos parámetros en el programa de conservación de la estirpe Guadyerbas de cerdos ibéricos.
Material y métodos
La población Guadyerbas fue establecida en 1945 a partir de 5 machos y 27 hembras que representaban el tipo negro lampińo de la vega del Guadiana y ha sido mantenida desde entonces como una piara cerrada. La gestión genética de la piara se llevó a cabo manteniendo una estructura familiar equilibrada y practicando apareamientos de mínimo parentesco (Rodrigańez et al., 1997). En el presente análisis se considera una genealogía integrada por 237 verracos y 751 cerdas agrupadas en 18 cohortes, correspondientes a animales nacidos en los sucesivos trienios del período 1945-97.
Para el análisis genealógico se han calculado para
cada cohorte las contribuciones, ![]() de cada
uno de los ancestros de la misma, definidas como la proporción de senderos que unen un
ancestro con un grupo de descendientes (James, 1962), así como el parentesco medio de
cada cohorte (incluyendo los recíprocos y el parentesco de los individuos consigo mismos).
Este parámetro puede expresarse en función de las contribuciones de los M
ancestros (de los que
de cada
uno de los ancestros de la misma, definidas como la proporción de senderos que unen un
ancestro con un grupo de descendientes (James, 1962), así como el parentesco medio de
cada cohorte (incluyendo los recíprocos y el parentesco de los individuos consigo mismos).
Este parámetro puede expresarse en función de las contribuciones de los M
ancestros (de los que ![]() son fundadores) de
la forma siguiente (Caballero y Toro, 1999),
son fundadores) de
la forma siguiente (Caballero y Toro, 1999),
![]() (1)
(1)
A partir de esta relación pueden definirse los siguientes parámetros:
Número efectivo de fundadores definido por Lacy
(1989) como el número de fundadores que, si contribuyeran de forma uniforme, producirían
la misma diversidad genética que la cohorte en estudio. Se calcula como ![]() siendo
siendo ![]() la contribución esperada de cada uno de los
la contribución esperada de cada uno de los ![]() fundadores.
fundadores.
Número equivalente de genomas fundadores. El parámetro anterior no tiene en cuenta que, incluso si los fundadores contribuyen igualmente a la cohorte, la variabilidad genética puede perderse por deriva genética. Por ello Lacy (1995) definió este nuevo parámetro como
![]() =
=![]() .
.
Numero efectivo de no fundadores definido por
Caballero y Toro (1999) como ![]() y que
refleja la contribución del resto de los individuos de la genealogía que no son
fundadores corregido para evitar redundancias debidas al parentesco entre los no
fundadores.
y que
refleja la contribución del resto de los individuos de la genealogía que no son
fundadores corregido para evitar redundancias debidas al parentesco entre los no
fundadores.
La relación entre los tres últimos parámetros es ![]()
Resultados y discusión
A partir de la relación (1) puede calcularse
para cada cohorte la contribución al parentesco de cada uno de los individuos fundadores.
Las contribuciones de cada fundador se estabilizan conforme avanza el tiempo. Sólo 14 de
ellos han sobrevivido hasta la última cohorte, extinguíendose la mayoría de los
restantes en la etapa inicial de la estirpe. El fundador que más contribuye al parentesco
de la última cohorte es la cerda Ruperta (1,61%) y el que menos Gévora
(0,00011%). La varianza de las contribuciones a largo plazo ![]() está directamente relacionada con el incremento asintótico del
coeficiente de consanguinidad. Así, en una genelogía con un censo constante por
generación N y un sistema de apareamiento regular es
está directamente relacionada con el incremento asintótico del
coeficiente de consanguinidad. Así, en una genelogía con un censo constante por
generación N y un sistema de apareamiento regular es ![]() (Wray y Thompson, 1980).
(Wray y Thompson, 1980).
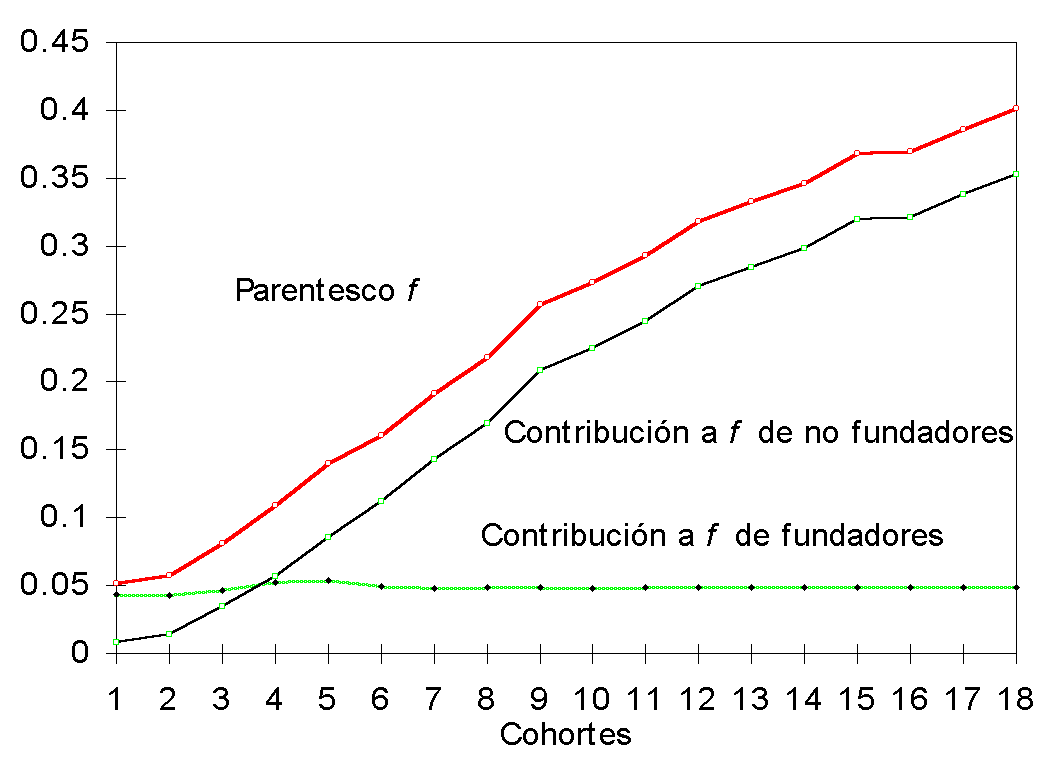
En la Figura 1 se presenta la evolución del
parentesco ![]() a lo
largo de los sucesivos trienios del período analizado, así como la contribución al
mismo de los animales fundadores y no fundadores. El valor de
a lo
largo de los sucesivos trienios del período analizado, así como la contribución al
mismo de los animales fundadores y no fundadores. El valor de ![]() en la última cohorte
es 0,401, siendo 0,048 y 0,353 la contribución respectiva de fundadores y no fundadores.
La contribución de los primeros se estabilizó en ese valor en el trienio 7, desde el que
el aumento del parentesco debe atribuirse a la contribución de los no fundadores.
en la última cohorte
es 0,401, siendo 0,048 y 0,353 la contribución respectiva de fundadores y no fundadores.
La contribución de los primeros se estabilizó en ese valor en el trienio 7, desde el que
el aumento del parentesco debe atribuirse a la contribución de los no fundadores.
Figura 1
Por otra parte en la Figura 2 se presenta la
evolución de los parámetros ![]() ,
,![]() y
y ![]() . Como se ha
explicado previamente
. Como se ha
explicado previamente ![]() se estabiliza con
el tiempo, mientras que el
se estabiliza con
el tiempo, mientras que el ![]() disminuye de
forma progresiva, desde un valor de 60 hasta un valor de 1,42 del primer al último
trienio, lo que refleja que la contribución al parentesco de los individuos no fundadores
va aumentando conforme transcurre el tiempo. Por último,
disminuye de
forma progresiva, desde un valor de 60 hasta un valor de 1,42 del primer al último
trienio, lo que refleja que la contribución al parentesco de los individuos no fundadores
va aumentando conforme transcurre el tiempo. Por último, ![]() , relacionado directamente con el coeficiente de parentesco también
disminuye como consecuencia del aumento progresivo de éste.
, relacionado directamente con el coeficiente de parentesco también
disminuye como consecuencia del aumento progresivo de éste.
Figura 2
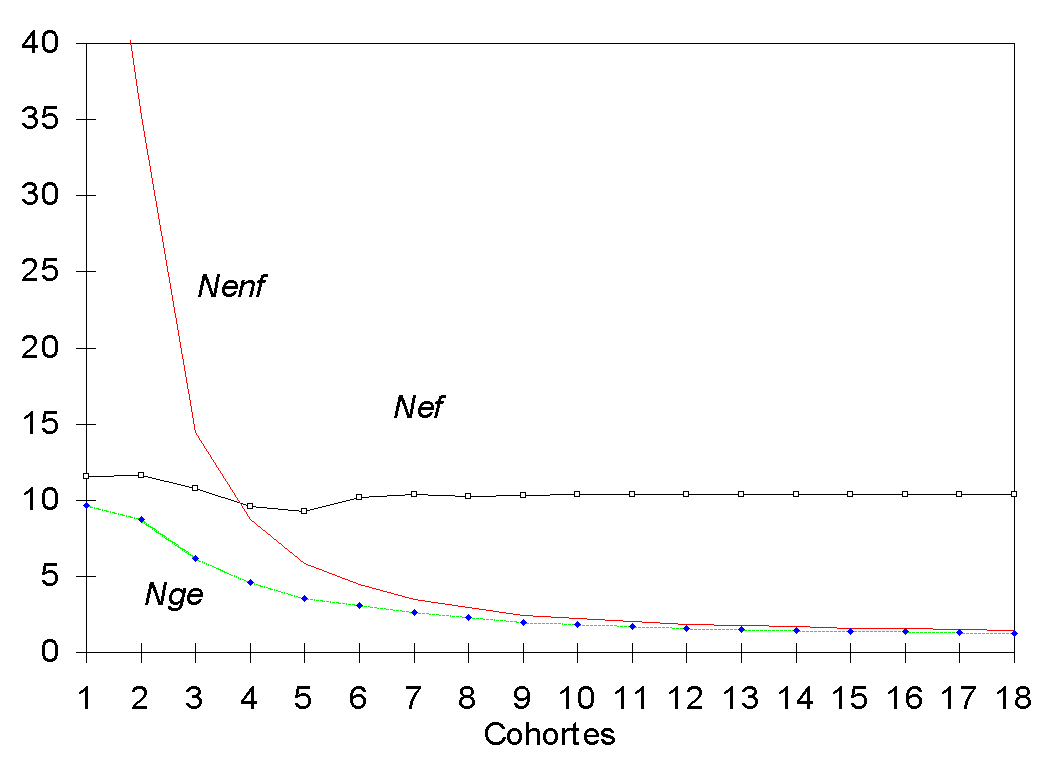
En génetica de poblaciones se ha utilizado, clásicamente, los conceptos de censo efectivo de consanguinidad (NeC) y censo efectivo de deriva (NeV) definidos como el censo de una población ideal que produjera la misma tasa de aumento de consanguinidad (D F) o la misma tasa de aumento de parentesco (D f) que la población considerada. Por otra parte, en la literatura conservacionista es más habitual el uso de los términos Nge, Nef y Nenf. En una población de censo constante y un sistema de apareamiento más o menos regular que garantice una aproximación asintótica, NeC= NeV, Nef= Ne/2 y Nge= Ne/t, siendo t el número de generaciones. Así, cuando el programa se estabiliza, Nef expresa la tasa de cambio de la consanguinidad por generación mientras que Nge, que disminuye de forma progresiva y continua, expresa la pérdida acumulada de variabilidad genética.
Otros parámetros utilizados en la literatura conservacionista, el número efectivo de ancestros y el de genomas fundadores, propuestos por Boichard et al. (1997), no parecen ser de interés ya que recurren a aproximaciones que no tienen en cuenta de forma correcta la deriva genética, subestimando la pérdida de genes. Ello se ha confirmado en este análisis, donde los valores calculados del número efectivo de ancestros y de genomas fundadores superan a los de Nge. Así para la última cohorte los respectivos valores son 6,81 y 1,38 frente al 1,25 obtenido por el procedimiento exacto aquí descrito.
REFERENCIAS
Boichard et al. 1997. Genetics Selection Evolution 29 : 5-23.
Caballero A. y M. Toro 1999. Genetical Research (enviado)
James J.W. 1962. Genetical Research 3 : 1-19.
Rodrigáńez et al. 1997. En International symposium on Mediterranean animal germplasm and future human challenges (Ed. D. Matassino, J. Boyazoglu y A. Cappuccio) pp : 183-186, EAAP Publication 85.
Wray N. y R. Thompson 1990. Genetical Research 55 : 41-54